L'évolution de la vie (3/3)
B) Les mécanismes de l'évolution.
Les mutations de l'A.D.N. au niveau des gamètes est
source d'évolution génétique, mais quelle est la
part de celle-ci dans l'évolution des êtres vivants ?
Comment expliquer aussi extinctions et radiations ?
1) Mécanismes d'innovation génétique.
Mutations :
Les mutations sont des modifications
affectant la séquence de l'A.D.N.. La fréquence de mutation
est faible pendant la réplication mais permet de créer de
nouveaux allèles pour un gène donné., ce qui accroît
la diversité. Certains agents dits mutagènes favorisent
l'apparition de mutations : rayonnement radioactif, produits chimiques … .
Mais même en présence de nombreux agents
mutagènes, les mutations sont insuffisantes à elles
seules pour expliquer la diversité du vivant car toutes les espèces
n'ont pas les mêmes gènes et justement les mutations ne permettent
pas de créer de nouveaux gènes.
Étudions le cas de l'hémoglobine humaine, formée
par l'association de quatre chaînes polypeptidiques identiques deux
à deux. Le type d'hémoglobine d'un individu dépend
de son âge :
- chez
l'embryon : 2 chaînes z
(chaîne du groupe A) et 2 chaînes e
(chaîne du groupe B) ;
- chez
l'embryon : 2 chaînes a
(groupe A) et 2 chaînes g
(groupe B) ;
- chez
l'enfant et l'adulte : de l'hémoglobine A – 2 chaînes a
(groupe A) et 2 chaînes b
(groupe B) –
mais
aussi un peu d'hémoglobine D – 2 chaînes a
(groupe A) et 2 chaînes d
(groupe B) –.
L'hémoglobine, qui sert au transport du dioxygène
dans le sang, se compose donc de chaînes du groupe A et d'autres
du groupe B. Les chaînes de ces deux groupes se ressemblent
mais les séquences géniques qui codent pour elles se trouvent
sur des chromosomes différents (A : chromosome 16, B :
chromosome 11). Les chaînes sont donc codées par des
gènes différents : tous les allèles d'un même
gène sont forcément sur le même locus mais il y a
6 loci différents pour 6 types de chaînes, il y
a donc 6 gènes.
Si les molécules produites se ressemblent tout en ayant des
fonctions identiques, c'est qu'elles sont homologues dans la même
espèce et le même individu. Ces molécules sont alors
apparentées et les gènes qui codent pour elles sont homologues
dans une même espèce. Ces gènes ne s'expriment pas
au même moment de la vie de l'individu mais existent simultanément
dans le génome : il s'agit d'une famille
multigénique. (-o zoom o-)
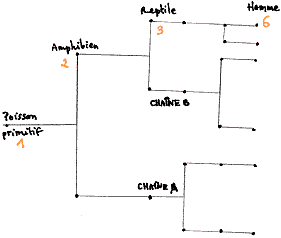
Ainsi l'hémoglobine existe chez tous les vertébrés
mais chez les agnathes et les requins (poissons primitifs), la synthèse
de l'hémoglobine est codée par un seul gène qui pourrait
être le gène ancestral. Chez les amphibiens, les gènes
sont au nombre de deux et chez les reptiles au nombre de trois. Il y a
donc apparition de nouveaux gènes avec l'évolution. L'homme,
mammifère, a hérité de trois gènes et en sont
apparus trois autres. On peut en déduire un arbre des gènes :
Création de gènes :
La création de gènes est possible par le mécanisme
de duplication-transposition : après
un chiasma mal effectué va avoir lieu un crossing-over inégal
où un chromosome reçoit un brin plus long que celui qu'il
a cédé. Les gènes présents sur ce brin mais
pas sur celui qui a été cédé se retrouveront
alors en double sur le chromosome … . Les mutations
permettront ensuite de différencier ce nouveau gène de son
original jusqu'à donner une famille multigénique si ce phénomène
est reproduit. Si la duplication-transposition n'est pas suivie
de mutations, il produit une famille multigénique de gènes
identiques (c'est le cas des gènes codant pour les histones). Mais
les mutations peuvent aussi entraîner un changement de fonction
des protéines produites ce qui rend nécessaire la consultation
des séquences pour s'assurer d'une parenté (c'est le cas
des gènes hypophysaires).
 |
Les mécanismes de duplication des gènes. Les
phénomènes génétiques à l'origine
de la duplication des gènes sont complexes. Parmi les mécanismes
possibles, les plus simples sont :
a. les cassures chromosomiques non homologues suivies
d'une réunion ;
b. la recombinaison homologue entre des éléments
répétés situés de part et d'autre du
gène.
Les deux copies sont rigoureusement identiques si l'évenement
survient chez un homozygote. Elles peuvent être légèrement
différentes si l'individu est hétérozygote.
|
Dans le cas des protéines de l'immunité, celles-ci
ont environ 30% d'acides aminés en commun. Les molécules
sont organisés en boucles, ces séquences d'acides aminés
sont homologues et on retrouve pour toutes les protéines de l'immunité
des domaines semblables, les gènes qui les codent sont donc apparentés.
On pense qu'il y a eu là aussi duplication-transposition qui a
conduit à la formation de plusieurs gènes codant chacun
un domaine. Après association de ces gènes (ou de fragments
de ceux-ci), cela a conduit au codage de sites complets : dans certains
cas il y a des domaines d'homologie issus des codes ancestraux, et dans
d'autres il y a des différences très fortes dues à
l'incorporation de gènes étrangers à la famille multigénique
(c'est le cas des molécules du C.M.H. qui présentent des
microglobulines assez différentes des immunoglobulines caractéristiques
de la famille). (-o zoom o-)
Le génome n'est donc pas figé, il y a au contraire
une extraordinaire plasticité du génome, indispensable à
l'évolution du vivant.
Création de nouveaux génotypes :
Les nouveaux génotypes obtenus doivent être
présents dans le génome des gamètes pour être
transmis. De plus il faut que l'innovation
ne soit pas fatale au porteur, c'est à dire qu'il soit compatible
avec la survie de l'individu et sa reproduction. Les mécanismes
de la reproduction sexuée (méiose et fécondation)
vont réaliser un brassage génétique et une transmission
aléatoire des allèles ce qui va faire que les descendants
auront des génotypes originaux.
2) Conservation de l'innovation génétique.
Innovation génétique et modification du phénotype :
Certaines innovations sont neutres, suite à des mutations
silencieuses principalement. Par contre d'autres ont des conséquences
énormes sur le phénotype. Ces mutations ont peut-être
servi à la modification des plans d'organisation lorsqu'elles ont
touché des gènes homéotiques / architectes.
(-o zoom o-)
Influence de la sélection naturelle :
Le cas exemplaire de la phalène
du bouleau nous renseigne sur les mécanismes de sélection
naturelle par rapport au milieu. En effet ce papillon est nocturne et
le jour il s'immobilise sur les troncs de bouleau. Comme les phalènes
sont généralement de couleur blanchâtre tachetée
de gris, une fois posés sur le tronc ils se confondent avec l'écorce
du bouleau. Il y a donc homochromie : l'espèce a une similitude
de couleur avec le milieu dans lequel elle réside, ce qui permet
par camouflage de se dissimuler d'éventuels prédateurs.
Pourtant avec la révolution industrielle (accompagnée de
pollution noircissant le tronc des arbres), un nouveau type de phalène
est apparue, cette fois-ci à la coloration noirâtre. Dans
les régions non-industrielles, les espèces claires sont
avantagées par rapport aux sombres, vites détectés
par les oiseaux. Dans les régions industrielles c'est le contraire
qui se produit.
La forme ancestrale est la forme claire mais en 1848 sont apparus
des mutants sombres. Par sélection naturelle
par le milieu, l'allèle favorisant la survie dans certaines
conditions sont transmis (allèle claire en milieu non-industrialisé …).
La fréquence d'un allèle dans une population dépend
ainsi du milieu (s'il est favorable ou pas dans celui-ci). La sélection
naturelle est donc un processus orienté qui favorise la survie
des individus dont le phénotype est le mieux adapté à
un moment donné aux conditions de l'environnement. Elle
modifie la fréquence allélique au sein d'une population
en augmentant celle des allèles présentant un avantage
sélectif.
Par ailleurs la sélection est dite conservatrice
si elle favorise le phénotype en place et novatrice
si elle tend à la production de mutants.
De même la répartition planétaire du gène
Hbs coïncide avec la distribution du paludisme par el fait que les
hétérozygotes HbA-Hbs présentent un avantage sélectif
qui entraîne une résistance à la malaria. La mutation
de l'hémoglobine est alors facteur de sélection naturelle
(bien que les homozygotes Hbs-Hbs soient atteints de trépanocytose).
Avant le rétablissement de l'isthme panaméen
les espèces étaient très différentes en Amérique
du Nord et du Sud. Mais cette voie terrestre ouverte, il y a eu des migrations
d'espèces du Nord vers le Sud et du Sud vers le Nord et le nombre
totale de familles a augmenté dans chacun des sous-continents.
Cependant, l'implantation des animaux venus du Sud n'a pas été
fructueuse contrairement à celle des espèces septentrionales.
Très vite le nombre de familles est revenu au même nombre
qu'avant les migrations. Certaines familles de migrants venus du Nord
ont pu se substituer aux espèces présentes en Amérique
du Sud. Il y a eu compétition entre
espèces (loup placentaire contre loup marsupial dans la même
espèce niche écologique) et seuls les plus forts ont survécu.
Il y a donc aussi sélection naturelle entre
espèces. (-o zoom o-)
Notion d'horloge moléculaire :
Le taux de mutation est à peu près égal
pour toutes les molécules mais les mutations ne sont pas conservées
au même titre, la vitesse d'évolution
dépend donc du taux de conservation des mutations. Ainsi
l'hémoglobine est une molécule essentielle et donc très
délicate à manipuler, il y a donc beaucoup de mutations
qui l'affecte qui produisent des allèles létaux ce qui diminue
fortement sa vitesse d'évolution (changement d'un acide aminé
tous les 5 Ma). Au contraire la fibrinopeptide est peu importante
pour l'organisme ce qui fait que les mutations peuvent être plus
souvent conservées sans pour autant compromettre la survie du porteur
(d'où une vitesse très élevée d'un acide aminé
changé tous les 1,25 Ma). Enfin dans le cas du cytochrome C,
enzyme intervenant dans les mécanismes respiratoires, la vitesse
d'évolution est extrêmement lente (changement d'un acide
aminé tous les 21 Ma) car la plupart des mutations qui l'affectent
sont éliminées avec le porteur.
Une fois que l'on connaît la vitesse d'évolution d'une
molécule on peut comparer cette molécule chez différentes
espèces pour retrouver la date possible d'existence de l'ancêtre
commun. Cela va permettre de préciser les arbres phylogénétiques,
c'est pourquoi on considère de telles molécules comme étant
des horloges moléculaires de l'évolution.
(-o zoom o-)
|